Cell Cycle Protein (CCP) Approach to In Situ Growth Rate Estimation:
Information on in situ growth rates (or cell division rates) is critical for understanding which of the processes, cell division (µ), import (I), grazing (G), cell death (M), or export (E), regulates the dynamics of phytoplankton populations.
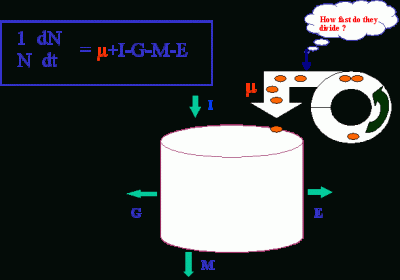
Practically, this information will be useful in assessing the contribution of a species (or a group of species) to carbon/nitrogen cycle, climate regulation, or harmful algal blooms.
In situ growth rate (µ) has been difficult to obtain due to lack of a feasible method. Over the past eight years, we have been developing a method named the cell cycle protein (CCP) method, which is based on the frequency of dividing cells technique (Weiler and Chisholm 1976, McDuff and Chisholm 1982) and the “terminal event” method (Carpenter and Chang 1988). All these share a common principle, that is the increase in population for the unicellular organism result from cell division. If the time sequential variation of the cell population that resides in a certain stage of the cell cycle can be determined along with the duration of this stage, µ can be estimated. The growth rate equation, developed for the “terminal event” method, is as follows,
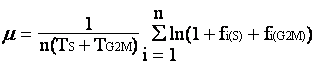
Basically, m is the growth rate (d-1), TS+TG2M is the duration of the terminal event, and fi(S)+fi(G2M) is the fraction of cells traveling through the S and G2-M phases in the ith sample. Advantages of this cell cycle approach are 1) no incubation of the organism is required; 2) growth rate estimates would not be affected by grazing, viral lysis, or physical transport of cells (assuming these processes are cell cycle independent).
There is a major improvement in the CCP method, which is the substitution of the presence of cell cycle proteins for the G2+M phase as the “terminal event” in the growth rate equation. Cell cycle proteins appear at specific stages in the cell division cycle and direct the cell to divide. Since they only appear in the cells that have reached a certain stage (e.g. G2+M phase of the cell cycle), their presence can be used to detect those cells. The presence of these proteins can be determined by using immunofluorescence on the whole cell with specific antibodies. The growth rate equation was modified (Lin et al. 1997) so that growth rate can be estimated regardless whether CCP-containing stages are terminal events. The equation now becomes:
eµ(Ts+TG2M) - eµTG2M - = 0
Where µ is the growth rate in d-1 to be resolved from this equation, TS and TG2M are the duration for the non-terminal event (represented by the S-phase) and the duration from this event to cell division (e.g. G2+M phase), respectively, and fS (bar) is the average fraction of a non-terminal event in the cell cycle over the diel sampling period. In the case that the cell cycle protein used represents a terminal event, i.e. TG2M = 0, the equation reduces to its earlier version (Eq. (1)). Alternatively, the expression of the CCP can be measured using reverse-transcription PCR, which is under investigation. The cell cycle proteins that have been explored include proliferating cell nuclear antigen (PCNA), p34cdc2, and cyclin B.